Dosen Pengampu: Putri Santika, S.ST.,M.Sc. Dewi F. Sabiku, S.P..M.Si. Dr. Ir. Rahmat Ali Syahban, M.Si. Teknisi: Prayitno S.P Rina Siofiana S.ST Anggota: 1. Siti Jamilatus Z (A41230468) 2. Wirdiyan Bagas K (A41230500) 3. Syaiyid Yusuf (A41230501) 4. Mirza Cahyo W (A41230518) 5. Ardi Firmansyah (A41230509) 6. Muhammad Sahar R (A41230526) 7. Tegar Firmansyah (A41230544) PROGRAM STUDI TEKNIK PRODUKSI BENIH JURUSAN PRODUKSI PERTANIAN POLITEKNIK NEGERI JEMBER 2024 BAB I PENDAHULUAN 1.1 Latar Belakang Benih merupakan salah satu faktor penentu keberhasilan dalam budidaya pertanian ter...
Dosen Pengampu: Putri Santika, S.ST.,M.Sc. Dewi F. Sabiku, S.P..M.Si. Dr. Ir. Rahmat Ali Syahban, M.Si. Teknisi: Prayitno S.P Rina Siofiana S.ST Anggota: 1. Siti Jamilatus Z (A41230468) 2. Wirdiyan Bagas K (A41230500) 3. Syaiyid Yusuf (A41230501) 4. Mirza Cahyo W (A41230518) 5. Ardi Firmansyah (A41230509) 6. Muhammad Sahar R (A41230526) 7. Tegar Firmansyah (A41230544) PROGRAM STUDI TEKNIK PRODUKSI BENIH JURUSAN PRODUKSI PERTANIAN POLITEKNIK NEGERI JEMBER 2024 BAB I PENDAHULUAN 1.1 Latar Belakang Benih yang bermutu dicirikan dengan tingginya viabilitas dan vigor benih. Sebelum...
Dosen Pengampu: Putri Santika, S.ST.,M.Sc. Dewi F. Sabiku, S.P..M.Si. Dr. Ir. Rahmat Ali Syahban, M.Si. Teknisi: Prayitno S.P Rina Siofiana S.ST Anggota: 1. Siti Jamilatus Z (A41230468) 2. Wirdiyan Bagas K (A41230500) 3. Syaiyid Yusuf (A41230501) 4. Mirza Cahyo W (A41230518) 5. Ardi Firmansyah (A41230509) 6. Muhammad Sahar R (A41230526) 7. Tegar Firmansyah (A41230544) PROGRAM STUDI TEKNIK PRODUKSI BENIH JURUSAN PRODUKSI PERTANIAN POLITEKNIK NEGERI JEMBER 2024 BAB I PENDAHULUAN 1.1 Latar Belakang Tanaman kopi merupakan tanaman tahunan yang mempunyai perakaranyang dangkal. Secara...
Putri Santika, S.ST.,M.Sc.
Dewi F. Sabiku, S.P..M.Si.
Dr. Ir. Rahmat Ali Syahban, M.Si.
Teknisi:
Prayitno S.P
Rina Siofiana S.ST
Anggota:
1. Siti Jamilatus Z (A41230468)
2. Wirdiyan Bagas K (A41230500)
3. Syaiyid Yusuf (A41230501)
4. Mirza Cahyo W (A41230518)
5. Ardi Firmansyah (A41230509)
6. Muhammad Sahar R (A41230526)
7. Tegar Firmansyah (A41230544)
PROGRAM STUDI TEKNIK PRODUKSI BENIH
JURUSAN PRODUKSI PERTANIAN
POLITEKNIK NEGERI JEMBER
2024
BAB I PENDAHULUAN
1.1 Latar Belakang
Dormansi merupakan benih adalah suatu keadaan ketika benih viabel tidak dapat berkecambah, meskipun berada dalam kondisi lingkungan yang optimum. Benih padi dikenal memiliki masa dormansi, yang disebut dengan istilah after ripening. After- ripening (AR) adalah proses pengaturan waktu dan lingkungan yang terjadi pada benih kering, yang menentukan potensi perkecambahan benih. Hal ini terindikasi disebabkan oleh adalah adanya sintesis hormon penghambat perkecambahan, yaitu ABA atau asam absisat.
Dormansi after ripening benih padi pada umumnya dapat patah dengan sendirinya setelah 8 minggu - 3 bulan setelah panen, apabila benih disimpan dalam keadaan kering. Namun apabila waktu menjadi hal yang tidak dapat ditoleransi, dan proses pengujian mutu benih ataupun penanaman tidak dapat ditunda, maka dapat dilakukan perlakuan-perlakuan khusus untuk dapat mematahkan dormansi after ripening pada benih padi, contohnya adalah dengan perlakuan air panas, dan KNO3.
Dormansi benih dipengaruhi secara morfologis, fisiologis, dan fisik benih (Baskin and Baskin, 2004; Finch-Savage and Leubner-Metzger, 2006). Dormansi benih dikategorikan menjadi dua penyebab, yaitu kulit dan embrio benih. Kulit benih dapat menyebabkan dormansi dengan cara mengganggu penyerapan air dan pertukaran gas, sebagai penghambat mekanis yang menghalangi inhibitor kimia keluar, begitu pula dengan embrio yang tidak berkembang sempurna (underdeveloped) dapat menyebabkan dormansi benih (Bewley and Black, 1985).
1.2 Tujuan
Untuk mengetahui metode pematahan dormansi benih padi yang efektif.
2Untuk mengetahui faktor faktor yang mempengaruhi dormansi.
BAB II TINJAUAN PUSTAKA
Dormansi benih merupakan atribut umum dari varietas benih padi dan ini mempersulit dalam memprediksi waktu kemunculan kecambah benih padi. Dormansi benih didefinisikan sebagai satu kondisi dimana benih tidak berkecambah walaupun pada kondisi lingkungan yang ideal untuk perkecambahan (Ilyas dan Diarni, 2007), dan merupakan karakter penting dalam program pemuliaan tanaman serealia karena berkorelasi dengan ketahanan benih untuk berkecambah sebelum panen (pre-harvest sprouting) sebagai faktor pembatas pada produksi padi di daerah beriklim lembab (Wan et al., 2006).
Sebagian besar benih padi memiliki sifat dorman yang sangat beragam dari 0 sampai 11 minggu sesudah panen (Ilyas dan Diarni, 2007). Dormansi benih padi secara alami akan berhenti setelah benih disimpan dalam kondisi kering yang disebut sebagai periode afterripening, Periode after-ripening esensial diperlukan yang memungkinan embrio benih mengatasi dormansi benih dan meningkatkan perkecambahan (Finch-Savage and LeubnerMetzger, 2006). Periode after-ripening berbeda antar spesies dan varietas, diduga disebabkan oleh keragaman genetik sifat dormansi antar spesies atau varietas tanaman (Carrera et al., 2008).
Dormansi benih dipengaruhi secara morfologis, fisiologis, dan fisik benih (Baskin and Baskin, 2004; Finch-Savage and Leubner-Metzger, 2006). Dormansi benih dikategorikan menjadi dua penyebab, yaitu kulit dan embrio benih. Kulit benih dapat menyebabkan dormansi dengan cara mengganggu penyerapan air dan pertukaran gas, sebagai penghambat mekanis yang menghalangi inhibitor kimia keluar, begitu pula dengan embrio yang tidak berkembang sempurna (underdeveloped) dapat menyebabkan dormansi benih (Bewley and Black, 1985).
Pemaparan benih dengan aplikasi eksogenus dapat meningkatkan perkecambahan dan mematahkan dormansi benih. Penelitian pematahan dormansi dengan pemanasan suhu tinggi dan perendaman larutan kimia (GA3, KNO3) berhasil meningkatkan perkecambahan 20 varietas unggul baru padi yang diuji (Wahyuni et al., 2011) dan 15 varietas unggul baru padi (Yuningsih dan Wahyuni, 2016).
Soejadi dan Nugraha (2002) menyatakan bahwa efektifitas metode pematahan dormansi sangat dipengaruhi oleh perilaku dormansi (intensitas, persistensi, dan mekanisme dormansi). Perilaku dormansi benih beragam antar genotipe atau varietas. Intensitas dormansi adalah persentase benih dorman pada saat panen. Semakin tinggi nilai intensitas dormansi mengindikasikan bahwa benih yang diuji memiliki tingkat perkecambahan yang rendah pada saat panen. Persistensi dormansi benih adalah periode simpan (dalam minggu) yang dibutuhkan benih dari saat panen sampai dimana persentase benih dorman telah mencapai ≤5% yang disimpan pada ruang simpan kamar (ambient storage).
Validitas hasil pengujian benih melalui pengujian daya berkecambah akan terganggu oleh dormansi benih. Bila pada akhir pengamatan diketahui masih ada benih dorman, maka pengujian harus diulang. ISTA Rules (2013) menyatakan apabila hasil pengujian daya berkecambah menunjukkan persentase benih dorman lebih besar dari 5%, maka pengujian perlu diulang dengan aplikasi pematahan dormansi.
BAB III METODOLOGI
3.1 Waktu dan Tempat
Praktikum Fisiologi Benih tentang Pematahan Dormansi After Ripening dilaksanakan pada hari Selasa, 29 Oktober 2024, Pukul 13.00-15.00 WIB. Di Laboratorium Teknologi Benih lantai 2, Produksi Pertanian, Politeknik Negeri Jember.
3.2 Alat dan Bahan
Alat
Germinator,
Baki,
Timbanagan,
Beaker glass,
ATK,
Kamera.
Bahan
Benih padi Baru panen,
Benih padi yang telah disimpan,
KN03,
Air,
Kertas merang,
Plastik,
Karet,
Kertas label.
3.3 Prosedur Kerja
Menyiapakan alat dan bahan ,
Menyiapkan masing-masing 100 benih padi untuk masing-masing perlakuan, sehingga diperlukan 300 butir benih padi baru panen dan 100 butir benih yang telah disimpan lebih dari 3 bulan,
Untuk perlakuan 3, menyiapkan air panas dengan suhu sekitar 50C, rendam sebanyak 100 benih baru panen dalam air tersebut selama 15 menit,
Untuk perlakuan 4, menyiapkan larutan 0,2% KNO3 sebanyak 200 ml, dan rendam media kertas merang dalam larutan tersebut,
Melembabkan kertas merang dengan air biasa, dan susun di atas plastic,
Untuk perlakuan 1,2 dan 4, susun benih di atas kertas merang yang telah dilembabkan dengan air biasa, tutup dengan kertas merang lagi, gulung, dan ikat,
Untuk perlakuan 3, susun benih di atas kertas merang yang telah dilembabkan dengan 0,2% KNO3, tutup dengan kertas merang lagi, gulung, dan ikat,
Beri label untuk masing-masing ulangan dan perlakuan, dan simpan di dalam germinator,
Melakukan pengamatan daya kecambah pada first count hari ke 7 dan final count hari ke 14.
BAB IV HASIL DAN PEMABAHASAN
4.1 Hasil
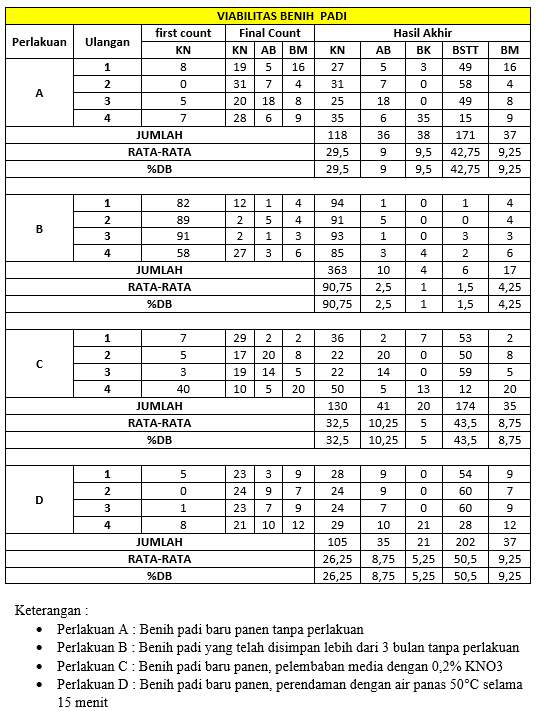
Berikut ini hasil daya berkecambah benih cabai berdasarkan berbagai pada berbagai tingkat kemasakan buah yang bias dilihat pada tabel 4.1 sebagai berikut.
Tabel 4.1 Hasil Penagamatan Laju Respirasi Kelompok A2.
4.2 Pembahasan
Pematahan dormansi benih padi melalui empat perlakuan yang memiliki hasil viabilitas berbeda. Perlakuan A (benih baru panen tanpa perlakuan) menunjukkan viabilitas dan daya kecambah yang rendah. Hal ini terjadi karena benih baru panen masih berada dalam fase dormansi, yang disebabkan oleh tingginya kadar hormon asam absisat (ABA) yang menghambat perkecambahan. Selain itu, kulit benih yang masih utuh menghalangi penyerapan air dan pertukaran gas yang diperlukan untuk memulai proses perkecambahan.
Perlakuan B (benih yang disimpan lebih dari 3 bulan tanpa perlakuan) menghasilkan viabilitas yang lebih tinggi dibandingkan perlakuan A. Hal ini disebabkan oleh proses alami after-ripening selama penyimpanan, di mana kadar ABA berkurang, sehingga embrio menjadi lebih siap untuk berkecambah. Proses ini juga dipengaruhi oleh kondisi lingkungan penyimpanan, seperti suhu dan kelembaban.
Pada Perlakuan C (pelembaban media dengan larutan 0,2% KNO3), viabilitas dan daya kecambah meningkat secara signifikan. Kalium nitrat (KNO3) membantu mematahkan dormansi dengan meningkatkan kemampuan benih untuk menyerap air melalui efek osmotik, yang mempercepat metabolisme benih. Selain itu, larutan KNO3 juga dapat mengurangi aktivitas inhibitor kimia yang menghambat perkecambahan.
Perlakuan D (perendaman dalam air panas 50°C selama 15 menit) menunjukkan hasil terbaik dalam meningkatkan daya kecambah. Air panas membantu melemahkan struktur fisik kulit benih, sehingga memungkinkan pertukaran gas dan penyerapan air yang lebih optimal. Suhu panas juga dapat menonaktifkan inhibitor kimia yang terdapat di kulit benih, seperti ABA, serta mempercepat proses aktivasi hormon pertumbuhan seperti giberelin. Proses ini memungkinkan oksigen masuk dan memfasilitasi pengeluaran karbon dioksida, yang merupakan bagian penting dari proses respirasi embrio selama perkecambahan. Selain itu, perendaman dengan suhu tertentu juga dapat menginaktivasi senyawa penghambat seperti asam absisat (ABA), yang sering terakumulasi di kulit benih. Dengan mengurangi pengaruh penghambat ini, embrio mendapatkan kondisi optimal untuk memulai pertumbuhan. Di sisi lain, Perlakuan juga menunjukkan daya kecambah tinggi, tetapi mekanismenya berbeda. Penyimpanan lebih dari 3 bulan memungkinkan terjadinya proses alami after-ripening, yaitu degradasi hormon ABA yang terjadi secara bertahap selama penyimpanan dalam kondisi kering. Hal ini membuat embrio lebih siap untuk berkecambah ketika kondisi lingkungan mendukung. Periode after-ripening juga memungkinkan waktu yang cukup untuk perbaikan internal benih, seperti pematangan embrio yang sebelumnya belum sempurna pada saat panen.
BAB V PENUTUP
5.1 Kesimpulan
Berdasarkan hasil pada tabel, terlihat bahwa metode pematahan dormansi benih padi memiliki efektivitas yang berbeda-beda. Perlakuan A (benih baru panen tanpa perlakuan) menunjukkan daya kecambah paling rendah karena benih masih berada dalam fase dormansi yang kuat. Perlakuan B (benih yang disimpan lebih dari 3 bulan tanpa perlakuan) menunjukkan peningkatan daya kecambah, yang disebabkan oleh proses after-ripening selama penyimpanan. Perlakuan C (benih baru panen dengan media larutan 0,2% KNO3) menunjukkan daya kecambah yang lebih baik dibandingkan Perlakuan A, karena KNO3 membantu mematahkan dormansi melalui peningkatan penyerapan air dan metabolisme benih. Perlakuan D (perendaman dalam air panas 50°C selama 15 menit) memberikan hasil terbaik, karena air panas efektif melemahkan kulit benih dan menonaktifkan penghambat kimia, mempercepat proses perkecambahan.
5.2 Saran
Praktikum ini sudah dilaksanakan dengan baik, dengan fasilitas yang lengkap dan pengelolaan waktu yang efisien. Namun, untuk meningkatkan kualitas praktikum selanjutnya, disarankan untuk menambahkan berbagai jenis bahan praktikum agar peserta dapat memperoleh wawasan yang lebih luas dan pengalaman yang lebih bervariasi. Selain itu, pengayaan materi dapat mendukung pemahaman yang lebih mendalam terhadap konsep yang dipelajari.
DAFTAR PUSTAKA
Baskin, J.M., Baskin, C.C., 2004. A classification system for seed dormancy. Seed Sci. Res. 14, 1–16.
Bewley, J.D., Black, M., 1985. Dormancy and the control of germination. In: Bewley, J.D., Black, M. (Eds.), Seeds: Physiology of Development and Germination. Plenum Press, New York, NY, pp. 175–233.Finch-Savage, W.E., Leubner-Metzger, G., 2006. Seed dormancy and the control of germination. New Phytol. 171, 501–523.
Carrera, E., Holman, T., Medhurst, A., Dietrich, D., Footitt, S., Theodoulou, F.L., Holdsworth, M.J., 2008. Seed after-ripening is a discrete developmental pathway associated with specific gene networks in Arabidopsis. Plant J. 53, 214–224.
Ilyas, S. dan W. T. Diarni. 2007. Persistensi dan pematahan dormansi benih pada beberapa varietas padi gogo. Jurnal Agrista 11 (2): 92-101.
Soejadi dan Nugraha, U.S. 2002. Karakterisasi sifat fisiologis benih plasma nutfah padi. Laporan Akhir Hasil Penelitian. Sukamandi: Balai Penelitian Tanaman Padi.
Wan, J. M., L. Jiang, J. Y. Tang, C. M. Wang., M .Y. Hou, W. Jing, L. X. Zhang. 2006. Genetic dissection of the seed dormancy trait in cultivated rice (Oryza sativa L.). Plant Scince 170: 786-792.
Wahyuni, S., Mira L. W., dan Rasam. 2011. Evaluasi karakteristik morfologis dan fisiologis benih serta metode efektif untuk pematahan dormansi benih beberapa varietas padi. Laporan Akhir Hasil Penelitian. Sukamandi: Balai Besar Penelitian Tanaman Padi.
Yuningsih, Aida F. V., S. Wahyuni. 2016. effective methods for dormancy breaking of 15 new-improved rice varieties to enhance the validity of germination test. Proceeding ISEPROLOCAL. Badan Penerbitan Fakultas Pertanian Universitas Bengkulu, Bengkulu, Indonesia, pp. 166-173.
Sepertinya ada yang salah dengan koneksi internet kamu. Harap sambungkan ke internet dan mulai menjelajah lagi.
AdBlock Detected!
Kami telah mendeteksi bahwa Anda menggunakan plugin pemblokiran iklan di browser Anda. Pendapatan yang kami peroleh dari iklan digunakan untuk mengelola situs web ini, kami meminta Anda untuk memasukkan situs web kami ke dalam daftar putih plugin pemblokiran iklan Anda.
Site is Blocked
Sorry! This site is not available in your country.